肠道菌群和氢气代谢密切相关
氢分子和甲酸(HCOO−)是哺乳动物肠道中许多初级发酵剂的代谢最终产物。两者在发酵中起着至关重要的作用,在缺乏外部电子受体的厌氧环境中,它们是单个微生物的电子汇。如果氢分子和/或甲酸盐在肠道生态系统中过多积累,初级发酵再生电子载体的能力可能受到抑制,微生物的代谢和生长中断。因此,消耗氢分子和/或甲酸的微生物,如产甲烷菌和同型醋酸菌,在维持初级发酵罐的代谢效率中发挥关键作用。为了宿主和环境的利益,人们对确定操纵哺乳动物肠道环境的方法越来越感兴趣。由于氢分子和甲酸盐是微生物物种间相互作用的重要媒介,了解它们的生产和利用可能是成功开发干预措施的重要切入点。反刍动物甲烷减排方法被讨论作为一个模型,以帮助理解氢分子和甲酸盐在肠道系统中的命运。
根据本综述提供的信息,我们对人类大肠内细菌产生氢气的相关代谢细节不了解,也缺乏具体理想的干预手段。虽然调整食物能影响肠道细菌产生氢气,也有一些研究表明这种方法增加产氢气具有全身效应。但这种方法是否可以作为长期干预手段仍然缺乏研究。肠道菌群和氢气的关系,通过肠道菌群调整氢气的补充方式都需要进一步研究。
有机异养厌氧微生物通过分解有机物发酵释放的单体和寡聚物获得能量。在缺氧环境中,这种发酵通常是由微生物之间的共营养相互作用所催化的。哺乳动物肠道生态系统中的厌氧环境是微生物栖息地,与非肠道系统如厌氧生物反应器(14-20天)和天然沉淀物(年或几十年)相比,它们有相对较短的食糜停留时间,每天转换一到两次。肠道系统的快速循环导致发酵不完全,主要产生挥发性脂肪酸(VFAs)、甲烷(甲烷)和二氧化碳(二氧化碳)。此外,还需要相对快速的ATP生成,这样微生物就可以生长得足够快,从而不会被从系统中过度清除。VFAs从肠道中被吸收,作为宿主动物的能量底物和代谢物,而甲烷和二氧化碳则作为气态废物排放。在反刍动物的前肠(瘤胃)和人的大肠中,甲烷和二氧化碳分别占可发酵碳的10%和17%左右,其余为VFAs[2,3]。相反,在生物消化器和沉积物中,VFAs也被代谢,有机物质被完全降解为甲烷和二氧化碳。
1981年,Meyer Wolin回顾了当时关于瘤胃发酵和人类大肠的知识。综述了氢分子和甲酸盐生产和利用在这些环境中的作用,以及不同微生物间相互作用在氢分子转移中的重要性。还讨论了减少甲烷产量以提高动物生产性能和饲料利用率的方法。从那时起,我们对肠道环境的认识迅速增长。虽然最重要的进展是确定了这些肠道环境中微生物种类的多样性,但在理解微生物及其基因组编码的酶的整体代谢方面也取得了进展。除种间氢分子转移外,种间甲酸转移也被认为是介导电子流的重要因素。此外,有研究表明肠道环境不是均匀的,而是由浮游和聚集的细胞组分(生物膜)组成,这些细胞组分与颗粒物质和肠道表面有不同的传代率,并且菌种间的电子流存在显著差异。近年来,研究的重点是了解肠道微生物群对人类健康的贡献,以及减轻反刍动物甲烷排放对环境的影响。然而,关于肠道微生物如何相互作用、如何与饮食成分以及它们的哺乳动物宿主相互作用,还有很多需要了解的地方。在这篇综述中,我们研究了反刍动物前前胃(瘤胃-网或瘤胃)和远端结肠的互惠发酵消化,强调了氢分子和甲酸的生产和利用在其代谢中发挥的作用。我们还讨论了可能改变氢分子和甲酸代谢并影响反刍动物甲烷生产的干预措施的进展。
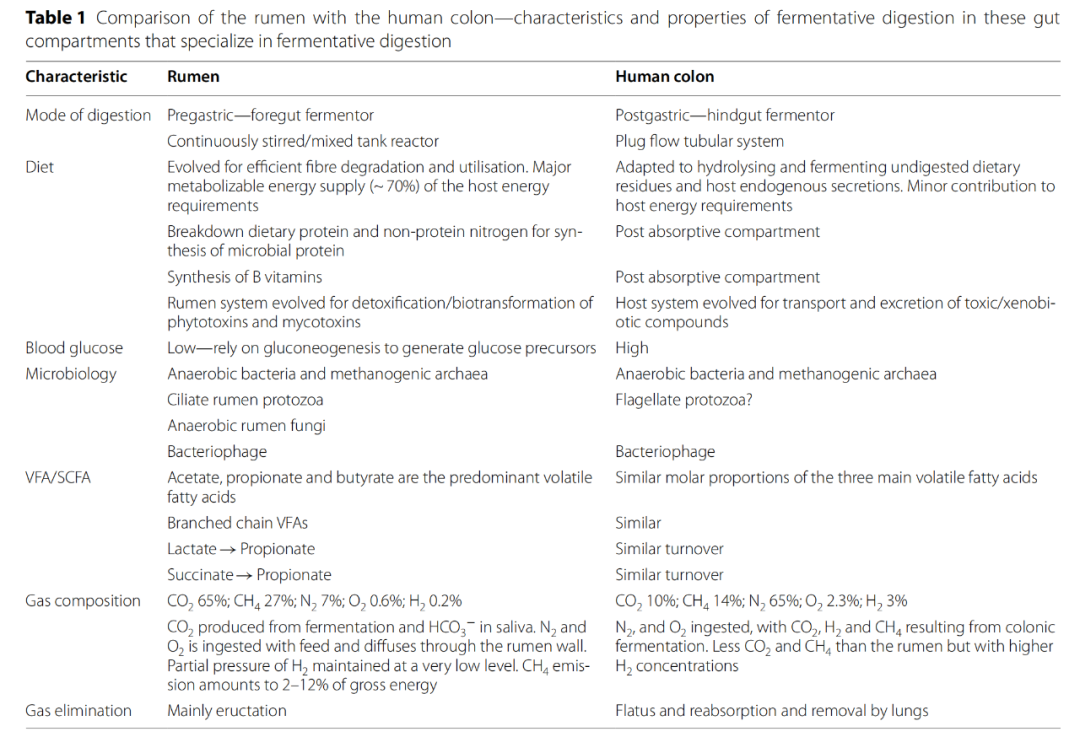
反刍动物的瘤胃是一个大的胃前发酵器官,在胃消化之前,微生物相互作用的发酵发生。相比之下,人类结肠是一个小得多的胃后发酵室。表1列出了这两种肠道发酵系统的重要特点。在对人类肠道的研究中,大多数样本是粪便或通过其他非侵入性技术获得的,而对于反刍动物,直接从插管动物的瘤胃或通过口腔插管收集食糜已被证实。此外,反刍动物的饲料成分和饲料添加剂的使用可以被精确控制,这使得科学实验比人类结肠的研究更加强大和严格。
这两个环境都有密集和多样的微生物种群,它们与宿主形成了紧密结合的生态单元。瘤胃中生活着多种微生物群落,包括厌氧细菌、产甲烷古菌、纤毛原生动物、厌氧植菌菌和噬菌体,众所周知,它们在代谢上具有很强的适应性,以应对饮食的变化。瘤胃与人类结肠的一个主要区别是存在大量的纤毛虫原生动物真核群体,占微生物生物量的50%。在正常的健康条件下,人的结肠主要容纳厌氧菌、产甲烷菌和噬菌体。在这两个系统中,微生物居民在其各自的宿主的营养、生理、免疫、保护和发育功能中发挥重要作用,但控制和塑造这些微生物群落的组成和活动的力量仍然知之甚少。
瘤胃和结肠之间的一个主要区别是,在瘤胃中微生物开始降解饲料,而在结肠中,宿主的消化过程首先对饲料起作用。养殖反刍动物的日粮主要由纤维(纤维素、半纤维素和果胶)和淀粉组成,其比例取决于生产系统,每天的摄入量相对稳定。人类的饮食变化很大,到达结肠的发酵底物包括未消化的膳食多糖,如纤维、抗性淀粉和逃避上消化道的低聚糖。宿主分泌的粘蛋白聚糖也是人类肠道微生物的重要底物。瘤胃微生物并不利用宿主聚糖,但某些瘤胃普雷沃菌中存在的宿主聚糖降解酶表明,它们可能能够利用唾液糖蛋白。
乙酸、丙酸和丁酸是瘤胃和人结肠发酵的主要VFA产物。瘤胃VFAs被吸收,贡献了70%的动物代谢能需求。VFAs也可从人体大肠吸收,有助于宿主的能量需求,尽管水平低得多(~ 10%)。一个重要的区别在于气态产物的产生。与瘤胃气体相比,人类肠道气体中二氧化碳和甲烷的比例较低,氢分子的比例较高。甲烷排放在瘤胃发酵中是普遍存在的,但人类甲烷排放者的比例存在差异,20%的西方人群被确定为甲烷高排放者。此外,氢分子很少是瘤胃发酵的最终产物,而是人体大肠发酵的主要产物,大量未被微生物利用的残留氢分子通过呼气或排气排出体外。
氢分子主要是在微生物发酵过程中由氢化酶产生的。这些酶催化碳水化合物发酵过程中被还原的辅助因子的再氧化,并通过还原质子来处理衍生电子生成氢分子。在瘤胃中,产甲烷古菌通过种间氢转移,将产生的大部分氢分子用于将二氧化碳或甲基化合物还原为甲烷。氢分子通过产甲烷作用保持在足够低的浓度,使发酵保持良好的热力学。
属于不同门的一系列瘤胃微生物已被证明可以产生氢分子, 65%的瘤胃细菌和古细菌基因组编码催化氢分子生产或消耗的酶。来自7种反刍动物胃肠道不同区域的宏基因组组合基因组(MAGs)得到了相似的结果。共检测到6152株[NiFe]-、[FeFe]-和含铁氢化酶的MAGs,其中3003株编码产氢酶(72.7%来自厚壁菌门),95株编码与氢营养产甲烷相关的氢分子-吸收氢化酶和甲基还原酶。
黄素电子分岔是一种电子对分裂机制,使能氧化还原反应与耗能电子转移反应耦合,可能对厌氧肠道环境中的发酵特别重要。对甲烷产量不同的绵羊进行的偏转录组分析表明,电子分叉的[fe]-氢化酶是瘤胃产氢的关键介质。碳水化合物发酵梭状芽胞杆菌(Ruminococcus, Christensenellaceae R-7组)的氢酶占所有氢酶转录本的一半,这表明这些微生物产生了氢营养甲烷短杆菌物种所使用的大部分氢分子。共培养实验表明,产氢纤维素发酵胃瘤菌在与产氢的富马酸还原菌Wolinella succinogenes共同生长时,表达其电子分叉氢化酶,并抑制其纯铁氧化还蛋白氢化酶。
瘤胃产甲烷菌还与通过其氢化酶体产生大量氢分子的原生动物存在共生关系。作为回报,原生动物受益于氢分子去除,因为高氢分子分压抑制了它们的代谢。原生动物消除研究的分析表明,去除纤毛虫原生动物可使甲烷的产量减少11%。产甲烷菌和厌氧瘤胃真菌之间也存在类似的关系,它们也含有氢化酶体。
宏转录组研究表明,虽然介导发酵氢分子生成的酶的表达水平相似,但产甲烷相关的转录本在高产绵羊中占主导地位,而在低产甲烷绵羊中,替代氢分子摄取途径显著上调。这些其他的氢分子吸收途径可能通过将氢分子的吸收从甲烷生成转向同乙酰生成(Blautia, Eubacterium)、延胡索酸盐和亚硝酸盐还原(Selenomonas, Wolinella)以及硫酸盐还原(Desulfovibrio)来限制甲烷的生成。同型产醋菌从氢分子和二氧化碳中产生乙酸,已知存在于瘤胃中,但它们的丰度一般低于氢营养产甲烷菌。在低氢分子浓度条件下,产甲烷菌在瘤胃中可能优于同型乙酸菌。硝酸盐和硫酸盐还原在热力学上比产甲烷和同生醋酸更有利,且硝酸盐和硫酸盐还原菌在瘤胃中自然存在。它们的种群密度随反刍动物饲粮中电子受体浓度的增加而增加。然而,反刍动物饲料中的硝酸盐和硫酸盐的浓度通常很低,因此这些过程的底物是有限的,它们的最终产物在高浓度时可能是有毒的。
关于甲酸盐在瘤胃中的浓度以及甲酸盐作为物种间电子载体的意义,我们所知甚少。在相对较高的氧化还原电位(较低的氢分子和甲酸盐浓度)下,甲酸盐在热力学和动力学上比氢分子更适合作为种间电子载体。这主要对浮游微生物是重要的,在那里,生物之间的电子转移距离大于那些生长在生物被膜和聚集物中的生物之间的距离。许多瘤胃微生物都含有甲酸脱氢酶基因,但与氢化酶基因相比,对它们在不同条件下的表达研究较少。
关于人体胃肠道中氢分子和甲酸盐的代谢机制,我们了解的较少。氢分子作为聚合碳水化合物发酵的最终产物,在结肠中大量形成,例如由厚壁菌门和拟杆菌门的成员。已知有两种处理氢分子的主要途径,即产甲烷和产乙酸,其中产乙酸是主要途径。当有硫酸盐时,硫酸盐还原菌进行硫酸盐的异化还原,但在没有硫酸盐的情况下,这些细菌通过产生氢分子而不是氧化它而茁壮成长。大多数产甲烷菌需要氢分子或甲酸盐才能生长,而同型醋酸菌具有多种代谢功能,也可以利用多种有机基质。一种途径对另一种途径的支配似乎在个体之间是不同的,并且是可以遗传的;在某些个体中,大部分生成的氢分子会转化为甲烷,而在另一些个体中,氢分子会积累到高浓度的。人类结肠食糜化学成分的空间和时间变化可能导致支持不同氢分子和/或甲酸代谢微生物的特定微生境。同相产醋在能量上不如产甲烷有利,但可以为宿主提供更大的好处,因为二氧化碳和氢分子可以转化为醋酸盐作为一种能源,而不产生气体。利用这两种途径的微生物代表了人类肠道微生物群的关键物种。
在纯培养条件下,氢分子和/或甲酸盐在某些结肠微生物中的形成和使用已经得到了很好的研究,但对氢酶和甲酸盐脱氢酶的研究还不够深入,而且我们对它们在发酵过程不同阶段的作用了解也比较少。只有一项研究调查了人类结肠中氢化酶编码基因的基因组和宏基因组分布,以推断氢分子循环的主导机制。来自人类微生物组计划胃肠道参考基因组数据库的大多数微生物物种都编码了形成或使用氢分子的遗传能力。各种各样的厌氧适应的氢化酶存在,以[fe]-氢化酶为主。对20名健康人粪便样本的宏基因组分析表明,所有样本的氢化酶基因含量主要由来自拟杆菌门(Bacteroidetes)和梭状芽胞杆菌门(clostridial)成员的发酵和电子分叉的[fe]-氢化酶组成。本研究认为,人结肠氢分子的代谢主要是通过发酵生成氢分子和利用电子分岔进行种间氢分子转移,而不是呼吸作用作为氢分子再氧化的主导机制。然而,这个粘膜生态系统的微环境龛位尚未得到充分的研究,氢分子和甲酸在人类结肠循环的细胞和遗传基础都需要进一步的探索。
属于克里斯滕森菌科的细菌是人类肠道微生物群中高度遗传的成员,与宿主健康状况有很强的相关性。对人类肠道宏基因组的分析已经确定克里斯滕森氏菌和史密斯甲烷短杆菌之间的积极联系。在共培养中,这些生物在密集的絮凝体中一起生长,克里斯滕森菌产生的氢分子支持史密斯氏菌产生的甲烷。这种相互作用使发酵最终产物倾向于更多的乙酸和更少的丁酸,潜在地影响人类宿主的生理。在对30个被鉴定为高或低甲烷排放源的对象的微生物群落分析中,smithii M. smithii和Christensenellaceae R-7组成员之间的相互作用也得到了强调。高排放源的特征是史密森氏菌数量增加1000倍,它与克里斯滕森菌科和Ruminococcaceae科的一个核心类群的关键物种的细菌同时发生。产生氢分子和甲酸的细菌和人类肠道同醋酸菌Blautia hydrogeneutrophica之间的交叉饲喂也在共培养中被证实
虽然人们有兴趣试图确定产甲烷菌的存在与人类肠道健康之间是否存在一种机制/因果关系,但开发能够实际减轻反刍动物甲烷排放的方法已经越来越紧迫。在全球范围内,反刍家畜肠道发酵产生的甲烷是农业温室气体的主要来源。理想情况下,任何已开发的缓解方法都应该为动物带来共同利益,例如提高生产或健康。协同效益可以帮助推动该技术在农场上的实际应用。虽然关于减少反刍动物甲烷排放的研究已经进行了多年,而且正在制定有希望的减缓方法,但这些方法并未出现一致的共同效益。这与经常提出的假设相反,即瘤胃产甲烷代表着能量的损失,从总能量摄入的2 - 12%,而这在原则上是可以用于动物生长或产乳的。历史上也有假说认为,抑制甲烷生成所导致的氢分子积累会损害纤维的消化和发酵。越来越明显的是,缺乏对氢分子和甲酸代谢及其操纵方式的了解是一个需要克服的障碍,以便支持开发具有共同效益的甲烷缓解方法。反刍动物中正在出现的甲烷缓解策略现在已经可用,可以提供模型系统,以帮助推进我们在这一领域的知识。下面讨论了四个有前途的领域。
动物的甲烷排放量各不相同,饲养甲烷排放量低的动物是缓解甲烷排放的一种方法。绵羊的研究取得了重大进展,研究发现动物产生的甲烷数量自然不同。该性状的遗传力使得低甲烷排放绵羊得以选育,选定的不同品系甲烷排放平均相差10-12%。饲料颗粒在瘤胃停留时间的缩短和瘤胃体积的减少等生理特性可能是导致甲烷排放较低的因素。瘤胃微生物群落也存在差异,低甲烷绵羊参与甲烷生产的微生物基因表达量降低。Kamke等人提出,低甲烷动物瘤胃微生物组支持异发酵生长,导致乳酸产生,乳酸随后主要代谢为丁酸盐。Greening提供了通过非产甲烷途径摄取氢分子的另一种解释,解释了观察到的差异。
在反刍动物的日粮中添加了几种替代电子受体,试图改变瘤胃发酵并降低甲烷的产量。硝酸盐是研究最多的化合物,它被亚硝酸盐还原成氨,降低了氢分子对甲烷合成的有效性。硫酸盐还原也将竞争电子和氢分子,并可能降低甲烷产量。通过抑制产甲烷菌来刺激醋酸菌的活性已被提出为反刍动物缓解甲烷的一种策略,但尚不清楚常驻瘤胃同型醋酸菌是否能够发挥处理氢分子的作用,或者是否需要将非常驻同型醋酸菌接种到瘤胃中。迄今为止的研究表明,有效的甲烷抑制剂(如3-硝基氧基丙醇)抑制了甲烷的生成,但并未证明同型乙酰生成增加。
在饮食与反刍动物相似的大足类有袋动物的肠道中,二氧化碳主要被还原为醋酸盐,而不是甲烷作为一种电子处理方式。大足类动物偏爱这一替代途径的原因尚不清楚,但有争论称,它们管状的前胃缺乏清除发酵气体产物的机制(例如,它们的免疫分泌物抑制负责释放氢分子或甲烷的微生物,以防止气体压力的累积,从而威胁肠道完整性。
几种不同的方法已经被用来专门针对瘤胃中的产甲烷菌。这些包括饲料添加剂,如3-硝基氧基丙醇、卤代化合物和某些海藻,以及开发抗甲烷菌疫苗的工作。迄今为止,现有技术的研究表明,当甲烷的生成受到抑制时,我们并没有观察到瘤胃氢分子排放的足够增加,以解释甲烷中没有捕获到的减少等量物。假设电子被转移到其他发酵产品,如醋酸盐,丙酸盐,丁酸盐,和微生物生物量,但这种电子重定向的平衡还不清楚。甲烷抑制剂与微生物的结合使用,可能会将氢分子导向其他产物,这还有待探索。
另一种方法是针对产生甲烷底物的微生物。尽管最近的工作已经开始确定最有可能产生氢分子和甲基化合物的细菌,这些化合物被用作瘤胃产甲烷的底物,但仍然存在显著的知识空白。在这一点上,还不知道减少底物产甲烷是否会阻碍整个发酵。
虽然单个动物产生的甲烷排放主要是由所食用饲料的数量引起的,但饲料的化学成分也会影响甲烷排放。因此,饲料消耗的性质可以选择具有不同发酵途径的微生物种群,它们产生的氢分子和甲烷更少。例如,以浓度为基础的饲粮与较低的甲烷产量(g/kg DMI),因为浓缩淀粉的发酵产生较多的丙酸和丁酸,而甲烷较少。发酵产物和微生物组成可能受到碳底物氧化状态的影响,特别是那些含有更多还原糖醇或更多氧化糖酸的产物。与多年生黑麦草相比,芸苔属牧草也被证明会导致羔羊甲烷产量降低。其原因尚不清楚,但已提出可能的原因是瘤胃微生物群的改变或芸苔属植物中存在具有生物活性的硫代葡萄糖苷。然而,一般情况下,反刍动物肠道甲烷排放的显著变化需要饮食的巨大改变。
在这里,我们对比了瘤胃和人类结肠环境,发现氢分子和甲酸盐是两种肠道生态系统中微生物群成员交叉进食的关键代谢物,在形成这些环境中运行的syntrophic网络中起着重要作用。氢分子和甲酸的产生作为单个微生物的电子收集器,并将电子转移到同生乙酸细菌和产甲烷古菌,这是确保反刍动物和人类持续的多糖降解和能量生成的关键功能。尽管厌氧菌和古菌在每种环境中大致相似,但瘤胃产生的氢分子主要是通过与甲烷结合来消耗的,而在人类结肠中,有大量的氢分子排放逃逸系统。确定哪些因素控制了这些差异,对于理解氢分子和甲酸代谢对人类肠道代谢的影响以及减少反刍动物甲烷对环境的影响具有重要意义。目前,新兴的缓解反刍动物甲烷的方法的可用性使瘤胃成为研究肠道系统中氢和甲酸盐的生产和利用的理想肠道系统,并产生适用于这两个系统的知识。
我们目前对哺乳动物肠道氢分子和甲酸盐经济的认识是不完整的,需要进一步了解参与氢分子和甲酸盐代谢的活性微生物群。模型氢营养菌,如甲烷短杆菌(Methanobrevibacter)和布劳提亚(Blautia)的培养和基因组序列是可用的,并已被用于演示种间氢分子和甲酸在共培养中的转移。然而,我们对哪些微生物在肠道环境中产生大量的氢分子和甲酸的知识是有限的。基于元基因组和元转录组的研究强调了编码氢营养营养特征基因的肠道微生物的多样性,并强调了对这些基因在肠道环境中功能的更精确信息的需要。目前还需要将更大比例的未培养微生物代表与其他宿主相关的同醋原和产甲烷菌菌株一起培养。这应该伴随着它们的生理、代谢和与其他肠道厌氧菌的相互作用的研究,以提供超出从基因组和宏基因组序列数据推断的知识体系。随着这种纯培养物及其相应基因组序列的可用性的增加,将证明有可能构建代谢相互作用的微生物组合,从而可以确定不同微生物对整体群落功能的贡献。
Kelly, W.J., Mackie, R.I., Attwood, G.T. et al. Hydrogen and formate production and utilisation in the rumen and the human colon. anim microbiome 4, 22 (2022). https://doi.org/10.1186/s42523-022-00174-z